El presente artículo es un resumen del publicado en La Cultura del Árbol núm. 72, titulado “Bases morfológicas para el análisis de la arquitectura de las plantas (I)“.
En este primer post se describen los procesos que rigen el desarrollo de las plantas.
Palabras clave: proceso de desarrollo, crecimiento, crecimiento determinado, crecimiento indeterminado, crecimiento rítmico, crecimiento continuo, preformación y neoformación, brote anual.
BASES MORFOLÓGICAS PARA EL ANÁLISIS DE LA ARQUITECTURA DE LAS PLANTAS (I).
1. PROCESO DE DESARROLLO.
El crecimiento primario de una planta es el resultado de múltiples procesos que se pueden agrupar en 2 distintos, pero coordinados, eventos morfogenéticos: organogénesis y extensión.
La creación de nuevos órganos (organogénesis) resulta del funcionamiento de células indiferenciadas que constituyen el meristemo apical (Fig. 1A). Localizado en el extremo del tallo este meristemo forma, cuando está en fase activa, pequeñas masas celulares, desarrolladas en hojas embrionarias, con diferentes potencialidades. La zona de inserción de la hoja en el tallo es denominada nudo y la porción del tronco que separa dos nudos sucesivos se denomina entrenudo.
Un tallo puede ser considerado como la entidad formada por la sucesión de entrenudos y como una entidad formada por el nudo, asociado a su hoja (u hojas) y yema/s axilares más el entrenudo subtenido, y representa la unidad estructural básica del cuerpo de una planta comúnmente llamado metámero o fitómero. Durante el crecimiento, la superposición y repetición de esta entidad elemental constituye el eje foliado (Fig. 1B).
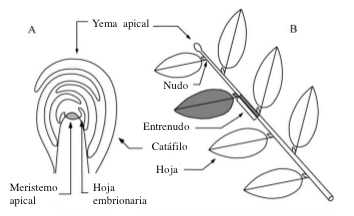
Fig. 1. Cada eje con hojas (B) termina en un meristemo terminal habitualmente protegido en una yema apical (A). Cada tallo consta de una sucesión de metámeros: el conjunto compuesto por (1) un entrenudo, (2) un nudo (el punto de inserción de las hojas en el tallo) localizado en su punta y (3) sus correspondientes hoja o varias hojas y yemas laterales asociadas (en gris en A).
1.1. Crecimiento determinado vs. indeterminado.
El ápice puede sufrir la abscisión o abortar después de algún periodo de funcionamiento o transformarse en una estructura especializada (flor, inflorescencia, espina, zarcillo, célula parenquimatosa, etc.) perdiendo la capacidad de extensión. En estos casos se considera que el eje tiene un crecimiento determinado (Fig. 2A-C). El crecimiento indeterminado se refiere a un eje donde su meristemo apical mantiene indefinidamente su potencial de crecimiento (Fig. 2D). Como el funcionamiento indefinido de un meristemo apical es siempre limitado, al menos, por la limitada vida útil de la planta a la que pertenece, este último término es algo “teórico” e indebidamente usado. Sin embargo, este concepto es útil y justificado para describir y nombrar este fenómeno.

Figura 2. El crecimiento determinado corresponde a una transformación irreversible del meristemo apical, que puede ser debida a (A) la floración apical como en Nerium oleander, (B) parenquimatización (flecha) del meristemo apical como en Alstonia sp. o (C) muerte apical o abscisión (“X”) como en Castanea sativa. El crecimiento indeterminado corresponde al funcionamiento permanente del meristemo apical, como se ilustra con el tallo principal de Picea excelsa (D).
1.2. Crecimiento rítmico vs. continuo.
Hallé et al. (1978) distingue entre ejes que no tienen marcado un cese endógeno de la extensión (crecimiento continuo) de brotes que sí que tienen marcado una periodicidad y cese endógeno de la extensión (crecimiento rítmico). Ambos patrones de desarrollo generalmente están más ligados a la extensión.
La extensión continua (Fig. 3A, B) es un fenómeno raro en el campo y generalmente ha sido observado y descrito en climas o ambientes ecuatoriales uniformes, principalmente en palmeras y manglares. En todos estos casos la producción de hojas y/o brotes es constante a lo largo del año. Cuando la extensión es continua, el eje resultante es generalmente bastante homogéneo y los sucesivos metámeros y sus elementos constituyentes son más o menos del mismo tipo y tamaño a lo largo del eje alargado (Fig. 3A, B, derecha).
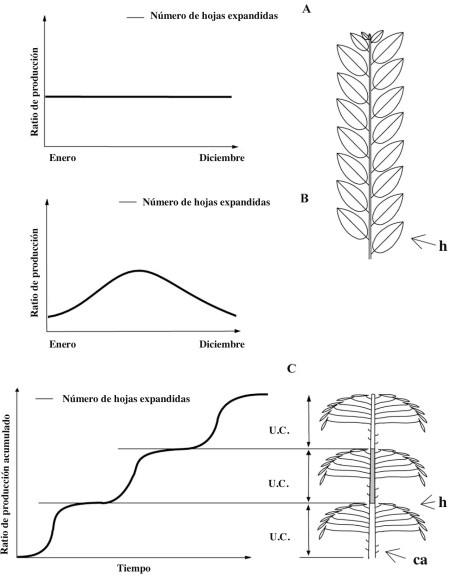
Fig. 3. Cada eje con hojas (B) termina en un meristemo terminal habitualmente protegido en una yema apical (A). Cada tallo consta de una sucesión de metámeros: el conjunto compuesto por (1) un entrenudo, (2) un nudo (el punto de inserción de las hojas en el tallo) localizado en su punta y (3) sus correspondientes hoja o varias hojas y yemas laterales asociadas (en gris en A). Fig. 1. Cada eje con hojas (B) termina en un meristemo terminal habitualmente protegido en una yema apical (A). Cada tallo consta de una sucesión de metámeros: el conjunto compuesto por (1) un entrenudo, (2) un nudo (el punto de inserción de las hojas en el tallo) localizado en su punta y (3) sus correspondientes hoja o varias hojas y yemas laterales asociadas (en gris en A).
La extensión rítmica se expresa mediante la alternancia de periodos de descanso de la actividad meristemática y periodos de extensión activa u “oleadas de crecimiento” (Fig. 3C). Hallé y Martin (1968), definieron “unidad de crecimiento” como la porción de un eje que se desarrolla durante un periodo de extensión ininterrumpido. Una unidad de crecimiento es generalmente fácil de identificar como el límite entre dos unidades de crecimiento que queda definida por una zona de entrenudos cortos y/o catáfilos (escamas foliares) correspondientes a los órganos protectores de la yema de la cual deriva (Fig. 4). Esta alternancia de catáfilos y hojas (Fig. 3C derecha, Fig. 4) puede no ser obvia (Fig. 5).
Lo más frecuente es que solo se conozca del crecimiento la componente de extensión, de tal forma que algunos autores han propuesto el uso del término “unidad de extensión” antes que unidad de crecimiento, aunque ambos términos son sinónimos.

Figura 4. Marcadores morfológicos de la extensión rítmica. Las fases de cese de crecimiento (flechas) y la delimitación de las sucesivas unidades de crecimiento (G.U.) como se revela a posteriori por una alternancia de catáfilas (ca) y hojas fotosintéticas (f) en Protea cynaroïdes (A) o sus cicatrices (Carya laciniosa, B, o Cicas pectinada, C).
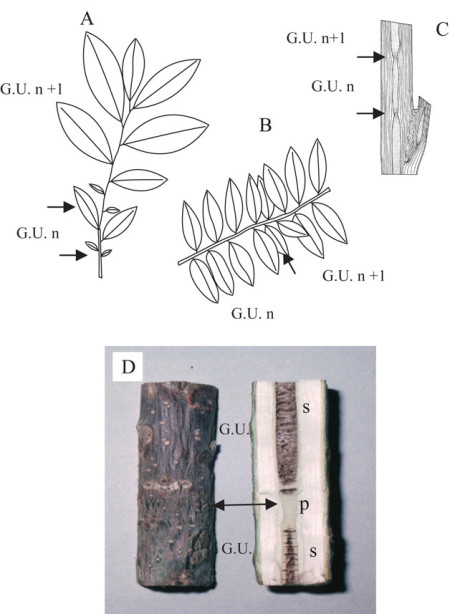
Sucesivas unidades de crecimiento pueden estar delimitadas (flechas) sólo por cambios más o menos marcados en el tamaño de la hoja (Virola michelii, A; Virola surinamentsis, B). En algunos casos el límite (flecha) entre dos unidades de crecimiento se muestra mediante una disminución del diámetro de la médula (Carapa procera, C) y/o incluso por la estructura de la médula como en Juglans sp. (D). G.U., unidad de crecimiento; “n”, “n+1”, sucesivos años teóricos de crecimiento; p, médula lisa; s, médula septada.
1.3. Preformación y neoformación.
En el caso del crecimiento rítmico, todos los metámeros y órganos del futuro tallo alargado pueden estar presentes en una fase embrionaria en una yema antes de la elongación, en este caso se denomina “preformado” y sus órganos constitutivos “órganos preformados” o “preformación”. Como consecuencia los tallos o brotes pueden comprender solo metámeros preformados o, más raramente, ser neoformados por completo. En muchos casos una parte preformada puede ser seguida por una neoformada, dando lugar a un brote mixto. La cantidad de preformación o extensión relativa de la preformación y neoformación en puede variar entre y dentro de las especies. Para ciertas especies arbóreas la preformación parece ser más relevante que la neoformación. Y las respuestas de neoformación en una posición específica de un árbol estarían implicadas en la respuesta plástica de los árboles a factores que actúan localmente en el momento de la extensión del brote.
1.4. Brote anual.
En los árboles de caucho que crecen en zonas ecuatoriales un eje con crecimiento indeterminado forma una nueva unidad de crecimiento aproximadamente cada 45 días, y estas unidades sucesivas son morfológicamente idénticas (Fig. 3C). En otras especies tropicales el tiempo de extensión rítmico es más complejo. En Ryania speciosa var. subuliflora el patrón de crecimiento anual corresponde a la extensión de dos unidades de crecimiento en un tiempo relativamente corto, seguido de una fase de reposo larga, seguida por un rápida emisión de dos unidades de crecimiento. En algunas especies de clima templado la extensión de los brotes puede ocurrir en uno, dos o más eventos sucesivos en una misma temporada dando lugar a una “brotación anual” formada por una sucesión de varias unidades de crecimiento o ciclos de crecimiento. Cuando se forman varias unidades de crecimiento sucesivas en el mismo ciclo vegetativo anual estas unidades de crecimiento suelen presentar características distintivas (Fig. 6); brotes de primavera y verano o brotes adicionales son distinguidos con frecuencia. En esta situación las unidades de crecimiento sucesivas producidas en un mismo año no son idénticas, y es útil y pertinente para distinguir el nivel de organización del brote anual.

La extensión del tallo puede producirse más de una vez durante el mismo año natural. El conjunto de las unidades de crecimiento producidas en un año se denominan brote anual (A.S.). En Quercus ilex (A) o Pinus halepensis (B) brotes bicíclicos, la primera unidad de crecimiento (G.U. 1) puede producir los órganos reproductivos mientras que la segunda (G.U. 2) es vegetativa. En tallos viejos, la presencia de conos femeninos o frutos en la primera unidad de crecimiento y el mayor desarrollo de las ramas nacidas en la segunda unidad de crecimiento de tales brotes anuales bicíclicos distingue a estas primera y segunda unidades de crecimiento, respectivamente, así como la delimitación a posteriori de las sucesivas brotaciones anuales (B). “n-1”, “n”, “n+1”, sucesivos años de crecimiento teóricos; flecha blanca sólida, límite de un brote anual; flecha blanca discontinua, límite de una unidad de crecimiento.
Referencias.
“Bases morfológicas y criterio para el análisis y descripción de la arquitectura de las plantas (parte I)”. Toni Serra Soliño. La Cultura del Árbol núm. 72, 2015. ISSN 1576-2777. Asociación Española de Arboricultura (AEA).
Barthélemy, D. & Caraglio, Y. 2007. Plant Architecture: A Dynamic, Mulitlevel and Comprehensive Approach to Plant Form, Structure and Ontogeny. Annal of Botany 99: 375-407. (www.aob.oxfordjournals.org)